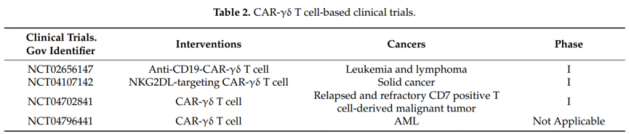
近期,National Cancer Institute (NCI)和Kite等联合发文-《Safety and feasibility of anti-CD19 CAR T cells with fullyhuman binding domains in patients with B-cell lymphoma》,介绍了新的CAR结构设计的全人源化的抗CD19 CAR-T细胞的I期临床试验(NCT02659943)。

图1 Hu19-CD828z和FMC63-28z的CAR结构设计图
具体CAR设计如图1所示,其中Hu19-CD828z是新设计,FMC63-28z是已上市的Yascarta的结构。这个试验结果显示,Hu19-CD828Z T细胞具有与表达FMC63-28Z的T细胞相似的临床抗淋巴瘤活性,55%患者获得了完全缓解。
然而,接受Hu19-CD828Z T细胞的患者中只有5%发生严重的神经系统毒性,而接受 FMC63-28Z T细胞的患者中有50%经历了这种程度的毒性(P=0.0017)。在接受Hu19-CD828Z T细胞的患者的血液中检测到的细胞因子水平低于接受FMC63-28Z T细胞的患者的血液,这可以解释与Hu19-CD828Z相关的较低水平的神经毒性。表达CAR的T细胞释放的细胞因子水平特别取决于CAR设计中包含的铰链和跨膜结构域。
由此案例出发,本文计划探讨工程化CAR的细胞外、间隔区、跨膜和细胞质结构域的最新进展,以及它们如何影响CAR-T功能。这种系统分析可以帮助揭示设计原则,这些原则可以广泛应用于未来的设计者免疫疗法。
1
CAR的基本结构介绍

图2 嵌合抗原受体结构概述
CAR结构包括三个主要部分胞外结合域、跨膜结构域和胞内结构域。胞外结构域由抗原结合域(通常是单链可变片段scfv)和铰链区构成。胞内结构域包括共刺激结构域和信号转导结构域构成。
以下分为胞外结构域、跨膜结构域和胞内结构域三部分来介绍。
2
胞外结构域
2.1
抗原结合域介绍
CAR胞外域设计的标准方法基于单克隆抗体,其可变区连接成单链可变片段或scFv(图3)。另一种在临床上用的方法是利用天然存在的受体-配体对。还有一种具有巨大潜力的相关方法是使用骆驼单域抗体片段,其特点是缺乏轻链,这种“纳米抗体”显示出有效的体内肿瘤靶向性,并且没有免疫原性。

图3 嵌合抗原受体 (CAR)-T 细胞的胞外结构域
scFv的四个重要特征是免疫原性、亲和力、特异性及其结合表位。
从鼠杂交瘤中获得的单克隆抗体 (mAb) 被发现在人类中具有免疫原性,导致低效力和立即从循环中消除,还表现出全身炎症反应,导致严重的生理并发症。因此,scFv的人源化有助于提高CAR的安全性和治疗潜力。
亲和力是一个关键参数,它解释了多种受体-配体相互作用,并受CAR表达水平、靶细胞上的配体密度和单个配体结合域的亲和力影响。亲和力已被调节以提CAR的特异性并减少“在靶点、肿瘤外”副作用,当靶标抗原在健康组织上普遍表达时,这一点尤其重要。亲和力调节可以影响CAR信号传导和其他效应器功能,如细胞因子分泌、增殖和持久性。
肿瘤相关抗原(TAA)是免疫治疗的主要靶点,在肿瘤组织中高度表达,在健康组织中也以较低水平表达。这会导致不必要的识别,有时还会导致危及生命的毒性。因此,在设计 CAR 以区分肿瘤细胞和正常组织时,scFv 选择至关重要。为避免这些并发症,应仔细了解 CAR scFv 与靶抗原之间的相互作用,并应在最终确定 scFv 之前进行交叉反应性研究。
目标表位的结构、空间可用性和功能方面也需要包括在CAR的设计考虑中。新的CAR设计,例如利用串联连接的scFv序列靶向两个抗原的双特异性CAR,可能需要额外的设计工作来确定合适的CAR结构,从而允许访问两个目标。
文献报道低亲和力的CD19 CAR与传统的基于FMC63的CAR相比,低亲和力抗CD19 CAR (CAT-CAR)显示出增加的抗原特异性增殖和增加的体内持久性,即使发现两者都靶向 CD19 抗原上的相似表位。两种CAR的IL-2和IFNγ分泌水平相当,而TNFα 在低亲和力 CAT-CAR(体外和体内的情况下略有增加)。

图4 FMC63 CAR 和CAT-CAR的亲和力差异
因此,scFv 的这四个方面(免疫原性、特异性、亲和力、目标定位)对于确定 CAR T 细胞的安全性和功能功效至关重要。
2.2
铰链区介绍
铰链/间隔区区域由CAR的非抗原结合细胞外区域组成。将scFv连接到跨膜域的间隔域为scFv提供了灵活性并有助于提高功效。已经观察到,铰链/间隔区通过提供柔韧性,延长长度,允许二聚化或改善稳定性来调节CAR功能。已经提出这些性质影响效应细胞-靶细胞相互作用,从而影响活化信号强度。
常见的铰链/间隔区已利用免疫球蛋白Fc,CD8a和CD28间隔区。当使用来自IgG1或IgG4的铰链-CH2-CH3间隔区构建CAR时,它们表现出与Fc γ受体的结合。这通过加速衰老、激活诱导细胞死亡等对持久性和活性产生负面影响,但CH2结构域的突变或缺失减轻了这些影响。
开头介绍的文章中作者同一个CAR将CD28铰链区换成CD8铰链区,就影响了神经毒性。
Almasbak等考察了没有铰链域的CAR-T细胞能否增强肿瘤杀伤是一个问题。因此,设计了两种有或没有铰链域的CAR来研究这个问题。最后,结论是铰链可以增强某些特定CAR-T细胞的扩增和抗肿瘤功效。因此,在设计CAR时应考虑抗原靶点位置和铰链长度,这对CAR-T细胞的活性至关重要。

图5 体内缺乏抗白血病活性与 IgG1-Fc 间隔区有关
为了保持最佳突触距离,膜远端表位通常需要较短的间隔区,而膜近端表位需要较长的间隔区。改变间隔长度会对CAR-T细胞的细胞溶解活性和信号传导产生深远的影响。
3
跨膜结构域介绍
CAR结构中的跨膜结构域作为将配体识别信号转导至细胞内胞质结构域的支点。多年来,CAR跨膜结构域的相关性在很大程度上被忽略了,因为这种结构成分被认为仍然是受体的惰性部分。跨膜区具有为CAR受体提供稳定性的能力,有助于与可能影响CAR T细胞功能的其他细胞表面蛋白缔合。铰链/间隔物设计的重要性差异很大,可能需要根据目标表位的相对位置逐案确定灵活性和距离要求。
为了阐明铰链和跨膜结构域在CAR 和 CAR-T 细胞功能中的作用,生成了不同的嵌合体并分析了它们在CAR-T细胞上的表达水平和抗原特异性活性。首先,创建了一个具有铰链、跨膜和信号转导域源自 CD3ζ,然后生成了六个 CAR 变体,其铰链或铰链/跨膜域源自CD4、CD8α和CD28。T细胞上的CAR表达水平和稳定性受跨膜而非铰链结构域的影响很大。大多数CAR-T细胞的抗原特异性功能取决于它们的CAR表达水平。
然而,尽管表达水平相同,但具有CD8α 或CD28衍生铰链域的CAR在CAR-T细胞功能方面表现出显着差异。这些结果表明[7],进入T细胞的CAR信号强度不仅受CAR表达水平的影响,还受铰链结构域的影响,铰链域调节CAR信号阈值,跨膜域通过控制CAR表达水平来调节CAR 信号量。

图6. 带有铰链域 (HD)或铰链/跨膜域 (HD/TMD) 修饰的血管内皮生长因子受体 2 (VEGFR2) 特异性嵌合抗原受体 (CAR) 的图示。
跨膜介导的相互作用也可用于产生新的CAR设计,例如可以传递跨细胞质域信号的拆分CAR系统。

图7. 跨膜域相互作用可以提供新颖的CAR设计[2]
4
胞内结构域
胞内结构域包括共刺激结构域和信号转导结构域构成。
4.1
CAR的不同代次
根据胞内共刺激域的差异,将CAR分为四个不同的代次。

图8 设计嵌合抗原受体 (CAR)-T 细胞:CAR 蛋白利用抗体的抗原识别能力
最基本的CAR设计,称为第一代CAR,包含一个抗原结合域,该域直接融合到TCR不变链CD3ζ的细胞内部分。第一代CAR通过磷酸化CD3ζ中基于免疫受体酪氨酸的激活基序 (ITAM) 域来启动激活级联反应。该信号不足以激活T细胞,需要外源性给予细胞因子。因此,最初的临床试验产生了令人失望的结果。
第二代CAR-T细胞将来自其他受体(如CD28、4-1BB (CD137))的共刺激结构域整合到CAR的细胞内片段中,以提高激活、增殖和存活率。这一概念在一项优雅的临床研究中得到了证明,将CD28作为共刺激掺入CD19 CAR-T细胞导致细胞扩增和存活率增加 [15]。
第三代CAR-T细胞结合了几个信号域,期望能增强 T 细胞的活化、增殖和存活。然而,在实践中,这种方法提供的临床益处仍在讨论中。
最近提出了第四代CAR-T细胞,因为它引入了一个细胞内结构域,能够触发由细胞因子诱导的信号(信号3)。
考虑到最终控制抗肿瘤功效的变量的多样性,每个特定共刺激信号的贡献对于每种肿瘤类型、胞外域和/或靶抗原是不同的。需要在CAR背景下对共刺激的分子免疫生物学进行系统的重新评估,以解决这些系统的复杂性,并为合理设计下一代CAR-T细胞制定指导方针。
4.2
共刺激结构域介绍
最常见的共刺激域是CD28和4-1BB。CD28共刺激提供更强烈的急性反应和有限的长期持久性的一般趋势,而4-1BB共刺激则相反。
已经研究了许多共刺激分子,包括CD28、4-1BB (CD137)、CD27、OX40 (CD134) 和 ICOS,它们已被纳入CAR的设计在临床中以进一步验证治疗效果。
多种共刺激结构域及其组合已在体外和体内进行了测试,但对于CAR-T细胞的理想设计尚未达成共识。共刺激域的选择可能取决于临床适应症以及其他 CAR模块。
Guedan及其同事探索了4-1BB、ICOS和CD3ζ的多种细胞质结构域组合,发现表达具有ICOS-4-1BB-CD3ζ细胞质结构域和ICOS跨膜结构域的CAR 的T细胞导致体内扩增和持久性增强35天内肿瘤消退100%。值得注意的是,具有ICOS–4–1BB–CD3ζ 但使用CD8跨膜结构域的CAR仅显示适度的肿瘤消退。鉴于这种CD8修饰的CAR也表达较低水平的IL-13和IL-6,作者认为细胞质结构域的近膜部分在确定CAR-T细胞的细胞因子谱中具有重要作用。

图9 包含与间皮素结合并在跨膜和细胞内结构域不同的 SS1 单链片段的嵌合受体的示意图

图10 ICOSBBz增强的体内功能仅在ICOS靠近细胞膜时观察到
4.3
信号转导结构域
ITAM是造血细胞受体胞质域中的序列基序。T细胞和传统CAR-T细胞在参与信号传导的ITAM数量方面存在主要差异。将ITAM的数量从三个减少到两个,可以选择性识别表达高目标密度的细胞。
马兹纳等人还表明,在基于4-1BB 的第二代CAR中,CD3ζ ITAMs 从3个增加到 6 个增加了T细胞增殖、IL-2 产生以及体内抗肿瘤反应。在另一项研究中,CD3ζ 的每个ITAM 都发生了突变,发现功能性ITAM的数量及其位置都会影响CAR-T细胞增殖、体内肿瘤根除以及分化表型。在同一项研究中,作者表明单个膜近端功能性ITAM与CD28细胞质结构域结合能够维持增殖、持久性和细胞毒性。

图11 野生型和突变型1928ζ CAR 的细胞质区域。
在CAR-T细胞中,受ITAM影响的信号机制尚不清楚。特别是,反式ITAM的可用性增强了 T 细胞中不同的信号机制。ITAM的这些方面对CAR设计有影响,对信号这些方面的更好理解可以极大地为 CAR-T 设计提供信息。
结论和挑战
CAR 结构的每个模块都独立和协同地影响 CAR 功能。
降低 scFv 的亲和力已被证明可以提高对肿瘤的选择性识别,从而降低对靶点和肿瘤外的毒性。然而,降低亲和力也会导致活化和抗肿瘤细胞毒性降低。选择正确的细胞质域可以增强这些信号。间隔区的长度和性质会影响免疫突触距离,并且对于确定细胞溶解至关重要。
与scFv 结合使用时,间隔区对于实现目标表位的可及性至关重要。跨膜结构域是关键的结构成分,有可能被用于具有共刺激结构域和 CD3ζ 结构域反式耦合的新型 CAR 系统。在这篇综述中,我们专注于讨论 CAR 分子本身的设计,但必须承认CAR蛋白以外的参数会显着影响CAR-T细胞功能也很重要。例如,转导载体系统的选择和CAR的插入位点对疗效有深远的影响。
许多研究采用合理的蛋白质设计和/或文库筛选来优化CAR序列。CAR组件可以根据其体外和体内功能以组合或顺序方式进行修饰。在一项研究中,使用两个不同的scFv序列靶向CD19和CD20的双特异性CAR依次确定了间隔区和 scFv接头要求,以实现对两种抗原的最佳靶向。
CAR-T 开发的第二个主要障碍是确定与体内功效相关的体外定量参数。已经表明,诸如体外活化和细胞因子释放等参数并不总是与所需的体内结果相关。最终,需要开发和验证用于CAR-T设计的体外功能分析,并与动物模型和临床数据的体内结果相关联。
沿着这条线,多功能性,即 T 细胞产生不止一种类型的细胞因子的能力,作为与体内功效相关的体外测量方法已经受到关注。使用一种新型的基于单细胞的测定法测试了输注前抗CD19 CAR-T 产品的32种不同细胞因子和趋化因子,据报道,增加的多功能性与改善的患者反应相关。可以定量询问预测体内 CAR-T 细胞功能的参数的功能分析平台将极大地促进相关 CAR 设计原理的阐明,能够更好地理解CAR-T细胞生物学,并促进 CAR 格式的发现新功能。
最终,这些努力可以简化免疫疗法的发现过程,从而以更低的成本和更短的开发时间获得有效的CAR-T候选药物。

参考文献
[1]Jennifer N.Brudno,Norris Lam ect. Safety and feasibility ofanti-CD19 CAR T cells with fully human binding domains in patients withB-cell lymphoma. Nature medicine. Doi:10.1038/s41591-019-0737-3.
[2]Albert t.Gacerez,Benjamine Arellano, and Charles L. sentman. How Chimeric AntigenReceptor Design Affects Adoptive T Cell Therapy. J. Cell. Physiol. 9999: 1–9,2016. Doi: 10.1002/jcp.25419
[3]Emiliano Roselli, Jeremy S. Frieling,Konrad Thorner, María C. Ramello, Conor C. Lynch,Daniel Abate?Daga. CAR?T Engineering: Optimizing Signal Transductionand Efector Mechanisms. Biodrug, 24 september 2019.10.1007/s40259-019-00384-z
[4] Sara Ghorashian, Anne Marijin Kramer ect. Enhanced CART cell expansion and prolonged persistence in pediatric patients with ALLtreated with a low-affinity CD19 CAR. Nature Medicine, VOL 25, September2019,1408–1414. doi: 10.1038/s41591-019-0549-5.
[5] Jayapriya Jayaramana, Michael P. Mellodya, Andrew J. Houb, RuchiP. Desaic, Audrey W. Fungd, An Huynh Thuy Phamd, Yvonne Y. Chenb,e,f , WeianZhao. CAR-T design: Elements and their synergistic function. EBioMedicine 58(2020)102931. Doi:10.1016/j.ebiom.2020.102931
[6] Almasbak H,Walseng E, Kristian A, Myhre MR, Suso EM, Munthe LA, Andersen JT, Wang MY,Kvalheim G, Gaudernack G, Kyte JA. Inclusion of an IgG1-Fc spacer abrogatesefficacy of CD19 CAR T cells in a xenograft mouse model. Gene Ther. 2015;22:391–403.
[7] Kento Fujiwara, Ayaka Tsunei, Hotaka Kusabuka,ErikaOgaki,Masashi Tachibana and Naoki Okada. Hinge and Transmembrane Domainsof Chimeric Antigen Receptor Regulate Receptor Expression and SignalingThreshold. Cells 2020, 9(5), 1182;
[8] Guedan S,Posey AD, Shaw C, Wing A, Da T, Patel PR, et al. Enhancing CAR T cellpersistence through ICOS and 4-1BB costimulation. JCI Insight 2018;3:1–17
[9] Feucht J, Sun J, Eyquem J, Ho YJ, Zhao Z, Leibold J, et al.Calibration of CAR activation potential directs alternative T cell fates andtherapeutic potency. Nat Med 2019;25:82–8.
[10] BrentjensRJ, Riviere I, Park JH, et al. Safety and persistence of adoptively transferredautologous CD19-targeted T cells in patients with relapsed or chemotherapyrefractory B-cell leukemias. Blood 2011;118:4817–28.
[11] Yu H,Sotillo E, Harrington C, et al. Repeated loss of target surface antigen afterimmunotherapy in primary mediastinal large B cell lymphoma. Am J Hematol2017;92:E11–E3.
[12] Blank C,Gajewski TF, Mackensen A. Interaction of PD-l1 on tumor cells with PD-1 ontumor-specific T cells as a mechanism of immune evasion: implications for tumorimmunotherapy. Cancer Immunol Immunother 2005; 54:307–14.
[13] Knepper TC, Saller J, Walko CM. Novel and expanded oncologydrug approvals of 2016-part1: new options in solid tumor management. Oncology(Williston Park) 2017;31:110–21.